�l���r�g��2024/10/8 9:23:29 ��x����77

������
������Ƥ������TEC���������ك����S֧���е��P�I������ͨ�^ճ�B�ԃ��к��N�������ӵ�ጷţ���T����ǰ�w�Ěw�����w�ƺͷֻ��������P�I���á����@��о��У����xС��TEC��ԭ�����B�����TEC-specific���wCK5��CK8�M���b�����@ЩTECͨ�^Գ�ﲡ����SV��40��T��ԭ�����D䛲����D������������Ȼ���҂����^��TEC��������TEC��iTEC���Ĺ��ܡ�ͨ�^�����@�R�zӰ�͂�����Y���Ϝy�����^����TECs��iTEC�ļ����ΑB����ֳ������Ȼ���M��ܛ��֬�y�����^�����¡�γ�����������IF��qPCR�z�yIL-7��Lptin��Pax-9��Sema3A����Ƥ�������P���ӵı��_ˮƽ����TEC�c�˼��Ԇκ˼�����Ѫ��������THP-1�������B������flow�����g��CFSE��ӛ�^��TECs�����MTHP-1��ֳ��Ӱ푡��y��˥�����P��b-������ø-dase�y���ԙz�y�����Ŀ�˥��������ͨ�^�⻯��ण�PI��Ⱦɫ�����������ڷֲ���ͨ�^Ĥ����V-PIȾɫ�z�yd��ɼ����PTX���T���ļ������������u�r�����Ŀ������������v�^���ģ��҂��l�F��������TEC��Ȼ������ԭ��TEC�������������B�����ܺ���Ƥ����;Ȼ����iTEC����ֳ�Ϳ�˥�Ϸ�����и������������о�������iTEC��SV40��T��ԭ�ɹ���������iTEC������W���������cԭʼTEC���ơ��@�N�������������������ٹ����о��đ�������ģ�ͣ���iTEC���ԭ��TEC��
����T�����l������Ҫ���٣������ڮa���������������m�������߹��ܵij���T�����������P�I���á�T�����܉��R�e������Ҫ�M�������ԏͺ��w(MHC)���ӣ������������M��ؓ�x����܉�^��������ԭ�ͷ�������ԭ��
�����Ƀɂ����܅^�ҽM�ɣ������|��Ƥ�ӡ�TEC�ǃɂ����g�е��P�I��Ԫ��TECs�Ͱl���е����ټ���֮�g��������Ǐ��s�����S��Ƥ�����W�j�l�����P�I�^�̣�Ҳ�����ټ����ڃɂ����م^�������l���еķֻ��ͳ�����P�I�^�̡��ڴ����g��TECs���ڵ����������P�I���á�����������W�J�R�������Mչ������TECs�ď��s���S�W�j�Լ����ټ�������Ƥ����֮�g������Ì��ھS�����������ٹ������P��Ҫ�����T�����l�����о���Ҫ�����ģ��ϵ�y�������������ɵ�������ȡ���ˣ������о������ژ������S����ģ�ͻ��˹����������(ATO)ϵ�y�������о����������T��������Ѫ�ɼ������漚��(HSPC)�е��x��ͳ��졣ͨ�^�����T������
Ȼ��������ȱ�����J��TEC����ϵ���@Щϵ�y�@ʾ���ܸߵČ��׃���ԡ����B��ԭ��TECs���w����Ɖ������ޣ��������ڼ���ˮƽ�ό����ٹ��ܵ��о���ȱ���������J�����ټ���ģ�͇����������ڼ���ˮƽ�ό����ټ�����TECs֮�g����õ��о�����ˣ�����һ�l������TECs��iTECs�����Ƿdz������x�ġ�
SV40T��һ�N�������ף��ɴ��M����ø�Ļ���錧���L���Ƅ���ʧ��[6]������о���������SV40T�����DȾ�������п���ʹ��ͬ��͵ļ������������������Ȯ���G����ϵ[7]���˸��Ǡ��ϵ[8]���i�Y�c��Ƥ����ϵ[9]�����о���Ŀ��������SV40T��������iTECģ�ͣ����^�켚�����Ժ��ܵ�׃��[10]���������ڞ��о����T�����l���ͻ��ڸɼ������w���T�������������ṩ�����Ĺ��ߡ�
ͨ�^���D䛲�������Դ����SV40T�D�Ƶ�TECs�С���4mg/mL��ù�غY�x���w�����B��Լ������^�쵽�ļ�����B���ã��ΑB������PCR�Y���@ʾ��iTECs�M��500bp������һ�l�������c�A�ڵ�Ŀ��Ƭ��λ��476bp��TECs��δ�@ʾ��Ŀ�ˎ�һ�£��D1A�����Y����������Դ����SV40T�ѳɹ����ϵ�TECs����M�С�TECs��iTECs�����ߟɹ�Ⱦɫ�@ʾ���������_��Ƥ�����خ��Ԙ�־��CK5��CK8��TECs��iTECs�ĵ��ͼ����|�ͺ�Ⱦɫ�Y����D2A��ʾ���������x�ļ����_����������Ƥ����������������������������ԭ��TECs����Ƥ���ԡ�
�Y���ό�����TECs��iTECs�M�ļ��������ڂ�����d1�r����ͬ�ġ�������d4��iTEC�M�ļ������@���࣬�@��ζ��iTECs����ֳ�����@����TECs���D1C�������⣬�ɷN�����ļ����ΑBҲ��ͬ��TECs��Ҏ�t�ęE�A�Σ������|�Ƕ̣����L�������ܣ���iTECs�����@�����Σ�������ɢ�Ҫ������L���D1����1D)���Y�������Toouli���˵ĽY��[17]��ܛ��֬�ϵļ����γɽY������TECs��iTECs�������γɼ������䡣����ֱ��
iTEC��200mmռiTEC�M�������59.52±14.14%����TEC�M�Hռ11.67±3.50%���D1B�����������ڽY���@ʾ��TECs��G0/G1��G2/M��S�����ı����քe��36.83±1.96%��18.43±5.81%��45.44±5.63%�������r��iTECs�ı�����27.99±2.52%��16.11±6.34%��54.40±8.0%���D3A�����Y����������������TECs���ΑB�l����׃����iTECs�ļ�����ֳ�Ϳ�¡�γ���������TECs��
�����u��TECs��iTECs��DNA�p���ķ����Ƿ�ͬ��ÿ������ϵ����¶��DNAǶ�넩PTX��ANNIXIN-V-FITC/PI�pȾ�Y���@ʾ��TECs-Control�M��TECs-PTX�M��iTECs-Control�M��iTECs-PTX�M���������ʷքe��16.70±3.70%��22.34±4.21%��9.92±2.79%��16.50±3.65%���D3B����ֵ��ʾ��ƽ��ֵ±�˜ʲn=3���Y������iTECs��PTX�T����DNA�p�������ܡ�����˥���ǙC�w�����[����һ�N�C�ƣ�Ҳ��˥�ϵ�ԭ��֮һ���ϻ��������F���c˥�����P��b-��������ø(SA-b-��������ø)Ⱦɫ�ʾGɫ��ԣ����@ʾ��ƽ�����ΑB[18]���Y����������TECs��iTECs���ڵ�3λ�r
������TECs��iTECs����Լ�����˥�ϼ������ı����քe��5.49%��5.81%���D3Ca��3Cc������TECs������6���r����Լ����ʞ�45.04%�����֮�£�iTECs�ڵ�12���r����Լ��������@�p�٣������ΑB�ʱ�ƽ��D3Cb��3Cd�����Y���������cTECs��ȣ�iTECs���и���ķ�����ֳ���������Ŀ�˥��������
�҂�ȫ��z�y��TECs��ֳ���ֻ��ͼ����^�����P�I���ӵı��_ˮƽ������TNF-a��IL-1b��IL-7��Sema-3A��Leptin��ACTH��FGFR2IIIb��Foxn1�������λ���Pax9.�붨��RT-PCR��qPCR���Y���@ʾTECs��iTECs���ڵ������������ӵı��_ˮƽ���ƣ���o�yӋ�W���x���D2B�����Y���C������������������Ƥ��������Ƥ���ܛ]�����@׃����
CFSEȾɫ��THP-1�����cTECs��iTECs�����B48h����ʽ�����x�z�yTHP-1������ֳ��r���Y���@ʾ�����սM��TECs�M��iTECs�MTHP-1���H���M�еı����քe��66.09±3.55%��54.51±7.18%��53.25±7.39%���cControl�M��ȣ�TEC�M��iTEC�Mĸ�w�����������ͣ���нyӋ�W���x��p<0.01����Ȼ����TEC�M��iTEC�M֮�g�IJ�]�нyӋ�W���x���D4A����
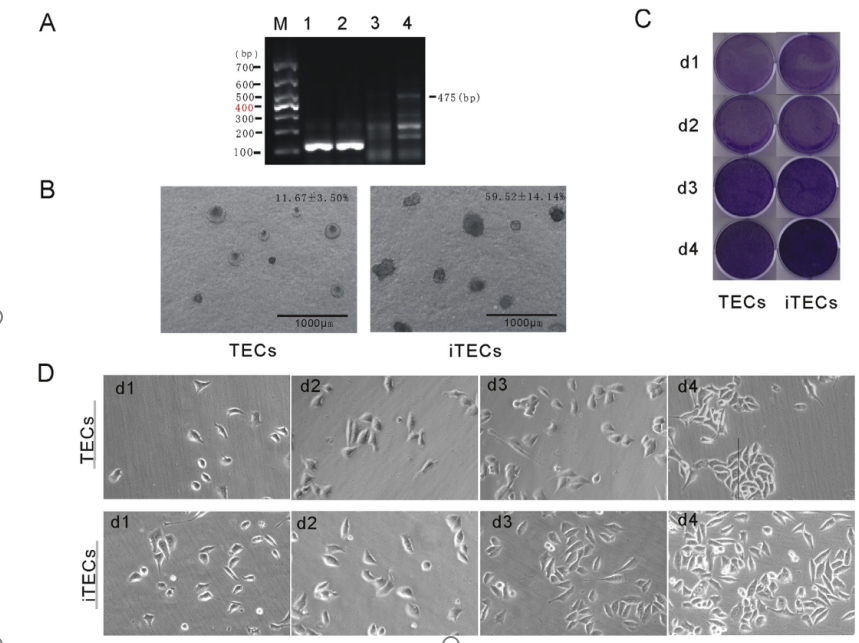
�D1.(A)PCRAssay�����z�Ӿ�z�yTECs��iTECs�е�SV40T��M��600bp���ΈD��TECs(1)��iTECs(2)��GAPDH��3��TECs(3)��iTECs(4)��SV40T��(B)TECs��iTECs��ܛ��֬�еľ������L������(C)�Y����Ⱦɫ���y��������������ֳ����������������ָ���r�g�c�ýY����Ⱦɫ��(D)��ͬͨ����ԭ����������С��������Ƥ�������ΑB�W�������P�ˈD�����ɫ�����Ľ�ጣ�Ո�x�߅������ĵľW�j�汾����
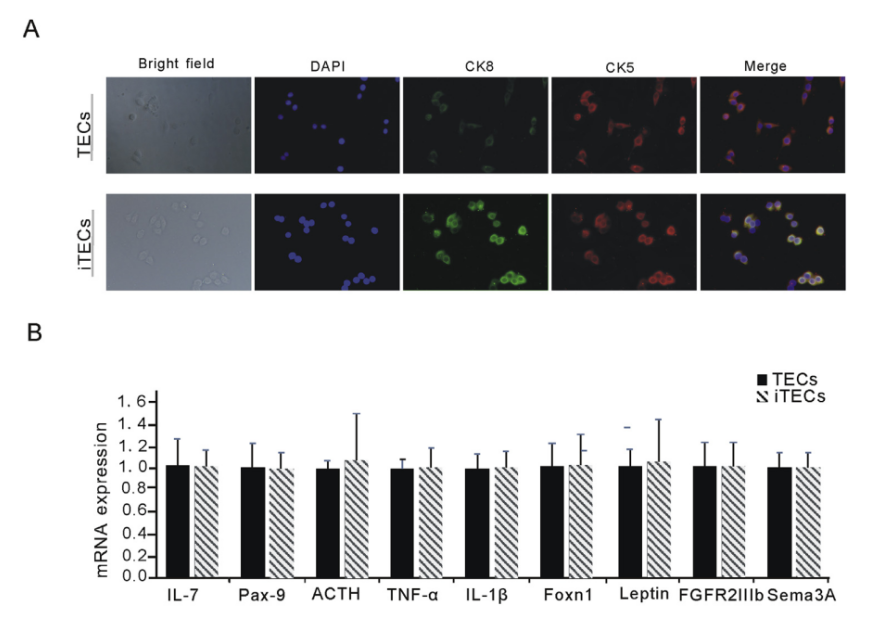
�D2.(A)iTECs���_�cTECs��ͬ����Ƥ�����خ��Ԙ�־�DAPI���{ɫ����CK5���tɫ����CK8���Gɫ���Լ�TECsGroup��iTECsGroup�ĺϲ��D����Ƭ���ڷŴ�400�����@�R���Ĕz�ġ�(B)�l�ΈD��ʾTECs��iTECs��TNF-a��IL-1b��IL-7��Sema3A��Leptin��ACTH��FGFR2IIIb��Foxn1��Pax9mRNAs���������_(ƽ��ֵ±S.E.M.)�������������������Č������P�ˈD�����ɫ�����Ľ�ጣ�Ո�x�߅������ĵľW�j�汾����

�D3.(A)TECs��iTECs�ļ��������ʡ�(B)PTX��TECs��iTECs�����ʵ�Ӱ푡�(C)�ڵ�3��(a)�͵�6��(b)��TECs�M�͵�3��(c)�͵�12��(d)��iTECs�M���M��˥�����P��b-galȾɫ����ɫ���^ָ���{ɫȾɫ�ļ�������
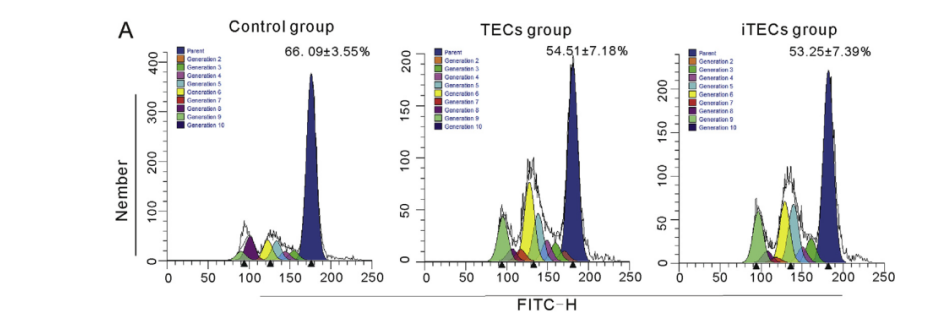
�D4.CSFEȾɫ���ڜyԇTECs��iTECs��THP-1������ֳ��Ӱ푡���������ĸ������(%)��ƽ��ֵ±SE,n=3��
�P�����ٹ��ܣ�TECs���о�����Ҫ�������֮һ����������ߌW���A�о��������˘O����Pע�������SV40T�錧�������������x��TECs�o���Ļ���������TECs�đ��þ�����Ҫ���x�����齨������������ϵ��õķ�������SV40���a�Ĵ�T��ԭ�ڸ�Ⱦ�����M����Դ���x�����S�����������D��������Ҫ���á����҂����о��У�SV40LT-SSR69�|�����Á���Ⱦԭ��TECs����ˣ���������TECs���������B��ײ��������B�m���f��iTECs���L���첢�����^�ߵ���ֳ�ʣ��D1C�������ϣ�iTEC���F���ѽ����˳��^45����������ֳ���á�iTECs�ļ����ΑB�Ȼ��������cTECs���ƣ�iTECs������Ƥ���������L������IF�Y���@ʾiTECs�����_CK5��CK8���Ķ�������TECs����Ƥ���ܡ�
�������ڷ�������������������ܛ��֬�����γɷ����Y��Ҳ������iTECs����ֳ�ʹ���ʃ���TECs��TECsS�ڼ�����45.44±5.63%������iTECs54.40±8.0%��TECsG0/G1�ڼ����ٷֱ��@������iTECs��PTX�T��������TECs�ĵ�����Ҳ����iTECs��TECs��iTECs��������ܛ��֬���B�������L�����oՓ��¡�Ĕ������С��Σ�iTECs�����и��@���Ŀ�¡�γ��������������������LTEC�����B�l���µļ����������҂��z�y���������־��“˥�����P��β-��������ø”��SA-b-gal����
�@���Է�������w���R�e��˥�ϼ�����β-��������ø��һ�N������ø���V���������w����w�Ȳ�ͬ��Դ��˥�ϼ�����[19]��˥�Ϻ�����������Ⱥ�еİ�������ø���Ա�������TECs��SA-b-galȾɫ�����������|��Ⱦ�ɾGɫ������ٷֱ��^�ߡ�
TECsͨ�^���ڲ�ͬ�Nļ������Ӂ��{��T�ܰͼ����������ؓ���x��TNF-α��һ�N��Ч�Լ������ӣ��������нM���ͱ��_[20]����������(IL)-7��T�ܰͼ����l�����w��ƽ��������ļ�������[21]������������1β(IL-1b)��Ҫ��Դ��������[2,22]���Sͻ�ų�����Sema3A���c�����е��S�༚���^�̣��������ټ�����ճ�����w��[23-25]���������ӣ������أ�������һ���̶���ͨ�^���،�T������ֱ�����Á��{���m��������[26,27]�����I����Ƥ�|����(ACTH)���{��������������������ã�������w�����B����[33,34])��Forkhead-box�D�����n1(Foxn1)���������䌦�л���9(Pax9)�{��TEC�ֻ�����δ������Ƥ�����ֻ��鹦����cTECs��mTECs[28-30]�����w�S�������L�������w2IIIb(FGFR2IIIb)��̖������������Ƥ��ֳ���P��Ҫ[31,32]�����@��о��У�������TEC��iTECs�ж�����ͬ��ˮƽ���_��
��֮��������TECs������W���Ժ����cԭʼTECs������W���Ժ��ܛ]���@������҂����о��������������������LTECs���w�����B�r�g��iMHCs�Ծ�������TECs�ķ��ڹ��ܣ��ɴ��M�κ˼�������ֳ�����о����о����ٺ����ټ����l�����w����ṩ����һ�N����ģ�͡�





�ߴa��ԃ�ſͷ�

�ߴa��ԃ�ſͷ�
©Copyright �Ϻ��ż�����Ƽ�����˾ All Rights Reserved. ICP�䰸̖����ICP��18032507̖-1 ���g֧�֣����x�W
�ſͷ�
��һ��������ԃ
�ſͷ�
��һ��������ԃ
021-34661275
021-34661276
15301693058
